1
FtsH1-11 | ATP-dependent zinc metalloprotease FtsH1-11
AS11 1789 | Clonality: Polyclonal | Host: Rabbit | Reactivity: A. acetabulum, A. thaliana, B. hypnoides (ulvophytes), G. theta, H. vulgare, Synechocystis sp., T. pseudonana, Z. mays
Benefits of using this antibody
- Product Info
-
Immunogen: KLH-conjugated peptide dereived from sequences of all known FtsH isoforms of Arabidopsis thaliana: FtsH1 Q39102 (At1g50250), FtsH2 O80860 (At2g30950), FtsH3 Q84WU8 (At2g29080), FtsH4 O80983 (At2g26140), FtsH5 Q9FH02 (At5g42270), FtsH6 Q1PDW5 (At5g15250), FtsH7 Q9SD67 (At3g47060), FtsH8 Q8W585 (At1g06430), FtsH9 Q9FIM2 (At5g58870), FtsH10 Q8VZI8 (At1g07510), FtsH11 Q9FGM0 (At5g53170) as well as 4 FtsH isoforms of Synechocystis sp. PCC6803NP_440330.1, NP_442160.1, NP_440797.1, NP_440525.1
Host: Rabbit Clonality: Polyclonal Purity: Serum Format: Lyophilized Quantity: 50 µl Reconstitution: For reconstitution add 50 µl of sterile water Storage: Store lyophilized/reconstituted at -20°C; once reconstituted make aliquots to avoid repeated freeze-thaw cycles. Please remember to spin the tubes briefly prior to opening them to avoid any losses that might occur from material adhering to the cap or sides of the tube. Tested applications: Blue Native PAGE (BN-PAGE), Western blot (WB) Recommended dilution: 1 : 500-1 : 2000 (WB) Expected | apparent MW: A. thaliana: FtsH1:76.7, FtsH2: 74, FtsH5: 75.2, FtsH6: 74.5, FtsH7: 87.8, FtsH8: 73, FtsH9:87.8 kDa (chloroplastic); FtsH3: 89.3, FtsH4: 77.2, FtsH10: 89.5 kDa, FtsH11:88.7 kDa (mitochondrial)
- Reactivity
-
Confirmed reactivity: Acetabularia acetabulum, Arabidopsis thaliana, Bryopsis hypnoides (ulvophytes), Guillardia theta, Hordeum vulgare, Synechocystis sp., Thalassiosira pseudonana, Thalassiosira punctigera, Zea mays Predicted reactivity: Diatoms including Phaeodactylum tricornutum, dicots, Chlamydomonas reinhardtii, Monocots; Nannochloropsis gaditana, Solanum lycopersicum
Species of your interest not listed? Contact usNot reactive in: No confirmed exceptions from predicted reactivity are currently known - Application Examples
-
Application example

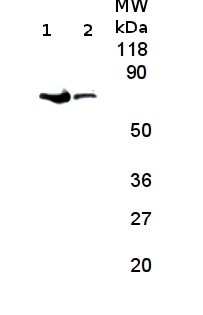
8 µg of total protein from Arabidopsis thaliana wild type (1) and FtsH 1 and 3 deletion mutant (2) extracted with 0.2M Tris-HCl pH6.8, 2%SDS, 10% mercaptoehanol, 5M Urea were separated on 12 % SDS-PAGE and blotted 2h to NC. Blots were blocked with non fat milk for 1h at room temperature (RT) with agitation. Blot was incubated in the primary antibody at a dilution of 1: 2 000 over night at 4°C with agitation. The antibody solution was decanted and the blot was rinsed briefly twice, then washed once for 5 min and 2 times for 10 min in TBS-T at RT with agitation. Blot was incubated in secondary antibody anti-rabbit IgG horse radish peroxidase conjugated) diluted to 1:10 000 in TBS-T+5% non fat milk for 2h at RT with agitation. The blot was washed as above and developed for 5 min with ECL according to the manufacturers instructions. Exposure time was 3 minutes.
Courtesy of Dr. Leah Nave, The Hebrew University, Israel
The dillution series of Hordeum vulgare (4; 2; 1 µg Chl) and Arabidopsis thaliana (3,6; 1,8; 0,9 µg Chl) thylakois were separated on 12% Criterion XT Bis-Tris SDS-PAGE (BioRad) gels and blotted for 25min 100V to PVDF membrane. Blot was blocked with 5% fat free skimmed milk in PBS-T for 1h at room temperature (RT) with agitation. Blot was incubated in the primary antibody at a dilution of 1: 500 overnight with agitation in 4˚C. The antibody solution was decanted and the blot was washed 3 times for 5 min in PBS-T at RT with agitation. Blot was incubated in secondary antibody (swine anti-rabbit IgG horse radish peroxidase conjugated) diluted to 1: 5000 in 1% fat free skimmed milk in PBS-T for 1h at RT with agitation. The blot was washed 5 min in PBS-T and 1 min in PBS developed for 5 min with ECL according to the manufacturers instructions. Exposure time was 90 seconds.
The band below FtsH2 is suspected to be FtsH5 as according to Sinvany-Villalobo et al. (2004): The most abundant FtsH transcript is that of FtsH2 (Fig. 3B), demonstrating a level similar to ClpC1. FtsH1 and 5 are also relatively abundant, each accumulating to levels of about 50% to 60% of FtsH2 transcript. Other FtsH transcripts are much less abundant, with FtsH6 not accumulating at all under optimal growth conditions.
Courtesy of Dr. Marta Powikrowska, University of Copenhagen, Danmark
- Additional Information
-
Additional information (application): For detection on a diatom samples, load of 5-10 µg/well is required as well as usage of enchanced chemiluminescence.The antibody also detects the chloroplast-encoded FtsH from Thalassiosira pseudonana (diatom) along with 3 related isoforms encode in the nuclear genome of Thalassiosira pseudonana.
For detection in BlueNative, recommened dilution is 1: 500. - Background
-
Background: FtsH belong to a family of ATP dependent peptidases. Localized in a chloroplast are following isoforms: FTSH1 (synonymes AAA, FTSH, FTSH Protease 1), Ftsh2 (VAR2, VARIEGATED 2), FtsH5 (VAR1, VARIEGATED 1), FtsH6 (FTSH PROTEASE 6), FtsH7, FtsH8. FtsH9.Localized in mitochondria are following isoforms: FtsH3 (FTSH Protease 3), FtsH4, FtsH10, FtsH11. - Product Citations
-
Selected references: Dogra et al. (2019). Oxidative post-translational modification of EXECUTER1 is required for singlet oxygen sensing in plastids. Nat Commun. 2019 Jun 27;10(1):2834. doi: 10.1038/s41467-019-10760-6.
Li et al. (2016). A Hard Day's Night: Diatoms Continue Recycling Photosystem II in the Dark. Front. Mar. Sci., 08 November 2016
Tietz et al. (2015). Functional Implications of Photosystem II Crystal Formation in Photosynthetic Membranes. J Biol Chem. 2015 Apr 20. pii: jbc.M114.619841.
Campbell et al. (2013). Photosystem II protein clearance and FtsH function in the diatom Thalassiosira pseudonana. Photosynth. Res. March 16. - Reviews:
-
This product doesn't have any reviews.
Accessories

AS09 602 | Clonality: Polyclonal | Host: Goat | Reactivity: Rabbit IgG (H&L)

AS05 094A | Clonality: Polyclonal | Host: Rabbit | Reactivity: Arabidopsis thaliana

AS11 1789S | Protein standard for quantitation