3
Anti-IDH | Isocitrate dehydrogenase (Cellular [compartment marker] of mitochondrial matrix)
AS06 203A | Clonality: Polyclonal | Host: Rabbit | Reactivity: A. thaliana, C.annuum, G. max, L. esculentum, O. sativa, P. sativum, S. tuberosum, Z. mays | cellular [compartment marker] of mitochondrial matrix
- Product Info
-
Immunogen: KLH-conjugated peptide 1 and peptide 2 conserved in all higher plants mitochondrial, NAD dependent isocitrate dehydrogenase subunits including Arabidopsis thaliana IDH-I Q8LFC0, At4g35260 and IDH-II P93032, At2g17130
Host: Rabbit Clonality: Polyclonal Purity: Immunogen affinity purified serum in PBS pH 7.4. Format: Lyophilized Quantity: 50 µg Reconstitution: For reconstitution add 50 µl of sterile water Storage: Store lyophilized/reconstituted at -20°C; once reconstituted make aliquots to avoid repeated freeze-thaw cycles. Please remember to spin the tubes briefly prior to opening them to avoid any losses that might occur from material adhering to the cap or sides of the tube. Tested applications: Immunofluorescence (IF), Western blot (WB) Recommended dilution: 1:400 (IF), 1 : 5 000 (WB) Expected | apparent MW: 39 | 45 kDa (Arabidopsis thaliana)
- Reactivity
-
Confirmed reactivity: Arabidopsis thaliana, Brassica oleracea, Capsicum annuum, Glycine max, Lycopersicum chilense, Nicotiana benthamiana, Oryza sativa, Solanum lycopersicum, Pisum sativum, Solanum sogarandium, Solanum tuberosum, Zea mays Predicted reactivity: Brachypodium distachyon, Brassica napus, Capsella rubella, Citrus sinensis, Cyanidioschyzon merolae, Galdieria sulphuraria, Hordeum vulgare, Malus x domestica, Medicago truncatula, Nicotiana tabacum, Phaseolus vulgaris, Theobroma cacao, Triticum aestivum, Vitis vinifera, Zea mays
Species of your interest not listed? Contact usNot reactive in: Chlamydomonas reinhardtii, Cyanophora paradoxa, Porphyridium purpureum
- Application Examples
-

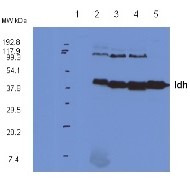
20 µg of total protein from Arabidopsis thaliana leaf extract (1), Arabidopsis thaliana fraction enriched with mitochondria (2), Arabidopsis thaliana pure mitochondria (3), Pisum sativum pure mitochondria (4), Solanum tuberosum pure mitochondria (5), were separated on 4-12% SDS-PAGE and blotted to nitrocellulose. Blots were blocked immediately following transfer in 5% milk powder in TBS. Blots were incubated in the primary antibody at a dilution of 1: 5 000 for 1h at room temperature with agitation, followed by an incubation with a secondary antibody and a series of washes. Blots were developed using chemiluminescent detection reagent.
* Band detected at ca. 90 kDa is suspected to be a dimmer of Idh, since this band is depleted upon peptide competition experiment.
Courtesy of Dr. Olivier Keech, Umeå Plant Science Centre, Sweden
15 µg of total protein stem extract from Lycopersicum esculentum (1), pure mitochondrial fraction isolated from stems of Lycopersicum esculentum (2), pure mitochondrial fraction isolated from stems of Capsicum annuum (3), pure mitochondrial fraction isolated from tubers of Solanum tuberosum (4) were separated on 10% SDS-PAGE and blotted onto nitrocellulose . After blocking with 5% milk in TBST , blots were incubated with the primary antibody at a dilution of 1:1000 in TBST for 1.5h at room temperature. Following incubation and wash steps, blots were incubated with secondary Anti-Rabbit IgG , Alkaline Phosphatase Conjugate for 1 hour at a dilution of 1:40 000. Blots were developed with the alkaline phosphatase detection system using NBT/BCIP.
Courtesy of Bartosz Szabala, Institute of Plant Genetics, Polish Academy of Science, Poland
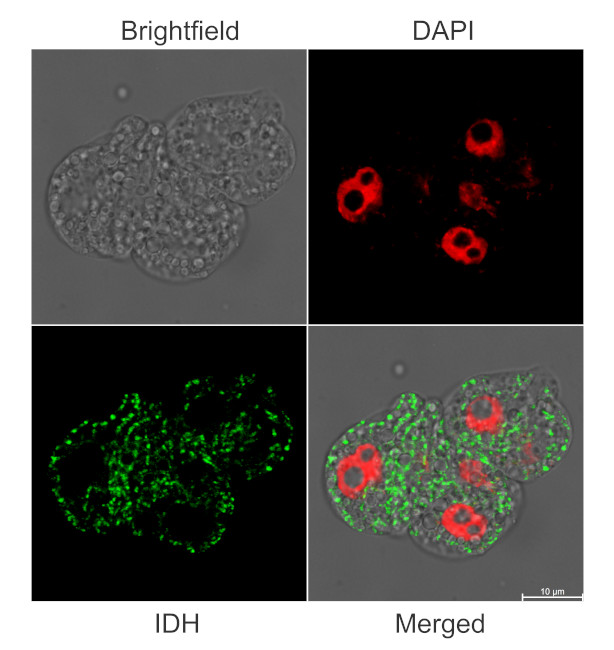
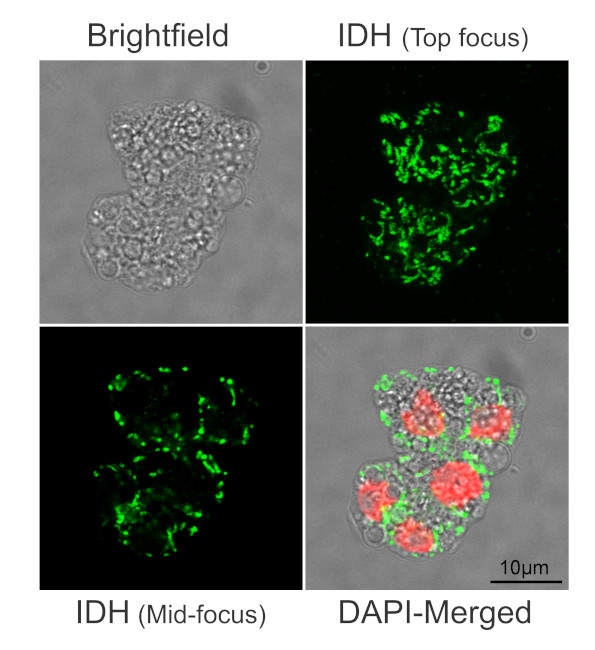
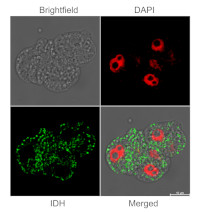
Immunofluorescent localization of IDH on suspension culture of Arabidopsis thaliana (upper image) or Oryza sativa (bottom image), using anti-IDH antibodies (AS06 203A) and anti-rabbit IgG DyLight®488 conjugated secondary antibodies (AS10 1165). DAPI staining of nuclei is pseudocolored red.
Material: Suspension cultures of Arabidopsis thaliana, ecotype Landsberg erecta cv.MM1 or Oryza sativa ssp.japonica cv. 'Unggi 9'
Fixation: Packed cell volume to fixer ratio: 250 µl : 5ml
Fixer composition and buffer: 4% (w/v) paraformaldehyde (freshly prepared as 8% stock and 0.2 µm filtered) in Phosphate Buffered Saline (PBS), pH 7.4 (2x stock, 0.2 µm filtered)
Container and method: in 6 cm Petri dish, gentle shaking at room temperature (RT)
Duration: 30 minutes (Arabidopsis thaliana) or 60 minutes (Oryza sativa). Cells were not shaken during the first 5 mins of fixation to allowed to partially recover from osmotic shock induced by formaldehyde.
Hydrophilization: no
Cell wall digestion: Yes
Packed cell volume to enzyme ratio: 100 µl : 2ml Enzyme composition: 1% (A) 1.2% (R) Cellulase (chromatically purified, powder, Worthington) 1% Pectinase (protease free, liquid, Sigma) Buffer: 0.5% (w/v) MES buffer, pH 5.6
Container and method: in 2 ml microfuge tube by rolling at room temperature (RT)
Duration: 30 minutes (Arabidopsis thaliana), 90 minutes (Oryza sativa)
Membrane permeabilization: Triton-X100 (0.2%), 10 min/RT
Antigen retrieval: no
Blocking buffer: Fish gelatin (5% v/v)
Washing buffer: PBS
Primary antibody dilution and incubation time: 1:400, ON/4ºC
Secondary antibody dilution and incubation time and supplier: anti-rabbit IgG DyLight®488 conjugated secondary antibodies (AS10 1165), 1:600, 1hn/RT
Co-staining of the nucleus (DAPI): Yes
Nucleus staining: 100 ng/ml DAPICourtesy of Dr. Ferhan Ayaydin, Hungarian Centre of Excellence for Molecular Medicine (HCEMM), Szeged, Hungary.

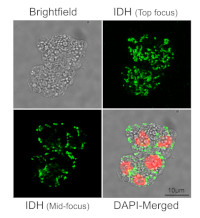
Sample: Maize protoplasts fixed in 4% paraformaldehyde in 1X PBS pH 7.4 for 1 hour and washed 3 times in 1XPBS buffer
Primary antibodies: Agrisera AS06 203A, anti-IDH
Secondary antibodies: Donkey anti-Mouse IgG (H&L), DyLight® 488 conjugated AS10 1201 (Agrisera)
Other reagents: 30% Bovine Serum Albumin solution, Sigma Aldrich Cat # A8577-50ML; Phosphate Buffered Saline 10X, Electron Microscopy Sciences Cat # 19342-10; Imaging spacer, Millipore Sigma Grace Bio-Labs SecureSeal™ imaging spacer Cat # GBL654008
Protocol: Block isolated protoplasts in a blocking buffer (2% BSA in 1x PBS) for 1 h at room temperature. Dilute the primary antibody to 1: 500 in a blocking buffer, mix well, and spin down at 150 x g for 3 min:
Spin down protoplasts in a blocking buffer at 150 g for 1 -3 min and remove as much supernatant as possible using a pipette tip with a cut end (wide opening). Resuspend protoplasts in diluted antibody and incubate for 1 h at room temperature.
Spin down protoplasts at 150 g for 1 min and remove as much supernatant as possible using a pipette tip with a cut end. Wash protoplasts in a blocking buffer 3 times for 5 minutes each.
Spin and resuspend as described in step 5 between each washing step.
Dilute the secondary antibody AS10 1201 to 1: 400, mix well, and spin down at 150 x g for 3 min:
Incubate protoplasts in diluted antibody for 1 h at room temperature.
Repeat the washing steps as described in 5-6.
Resuspend a protoplast pellet in a small volume of PBS 1X, pH 7.4.
Prepare microscopy slide: attach imaging spacer on top of the slide, peal off top adhesive membrane, add protoplast suspension on to the slide inside the spacer opening, cover with 22x22 mm cover glass No 1.5.
Imaging: Zeiss Elyra 7 SIM
Courtesy of Dr. Anastasiya Klebanovych, Dr. Kirk Czymmek, Dr. Kevin Cox, Dr. Blake Meyers, at the Donald Danforth Plant Science Center, USAApplication examples: 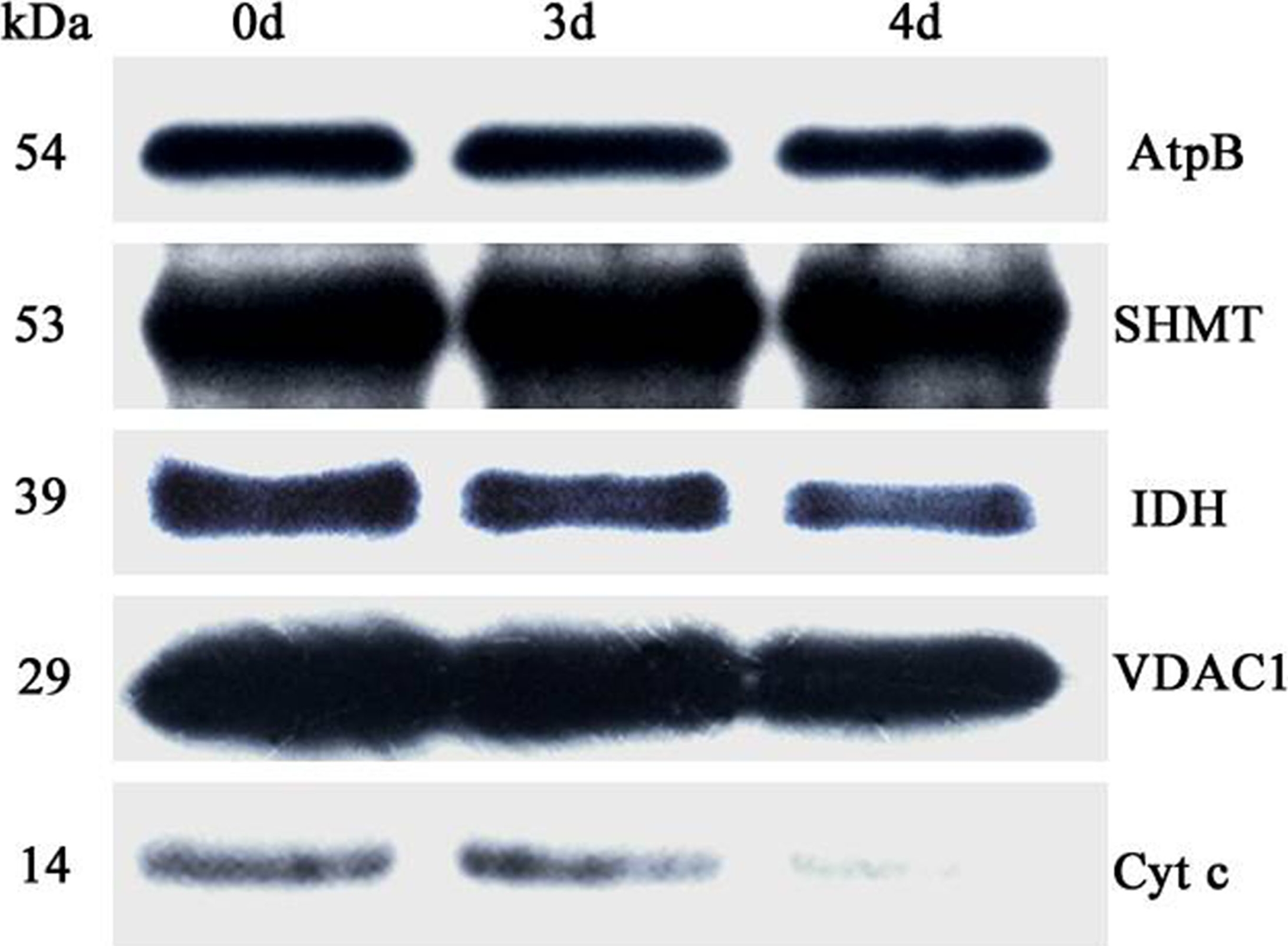
Reactant: Oryza sativa (Asian rice)
Application: Western Blotting
Pudmed ID: 27124767
Journal: PLoS One
Figure Number: 6A
Published Date: 2016-04-29
First Author: Yin, G., Whelan, J., et al.
Impact Factor: 2.942
Open PublicationAbundance of a variety of known mitochondrial proteins by the immunoblot analysis of mitochondria isolated from 0 d, 3 d, and 4 d aged rice embryos after 48 h imbibition.Total 10 ?g protein was separated by SDS gel electrophoresis and blotted to supported polyvinylidene difluoride, then probed with antibodies against beta subunit of ATP synthase (AtpB), serine hydroxymethyltransferase (SHMT), isocitrate dehydrogenase (IDH), voltage dependent anion channel 1 (VDAC1) and cytochrome c (Cyt c).
Reactant: Nicotiana tabacum (Common tobacco)
Application: Western Blotting
Pudmed ID: 31985876
Journal: Plant Biotechnol J
Figure Number: 3C
Published Date: 2020-09-01
First Author: Eseverri, Á., López-Torrejón, G., et al.
Impact Factor: 8.66
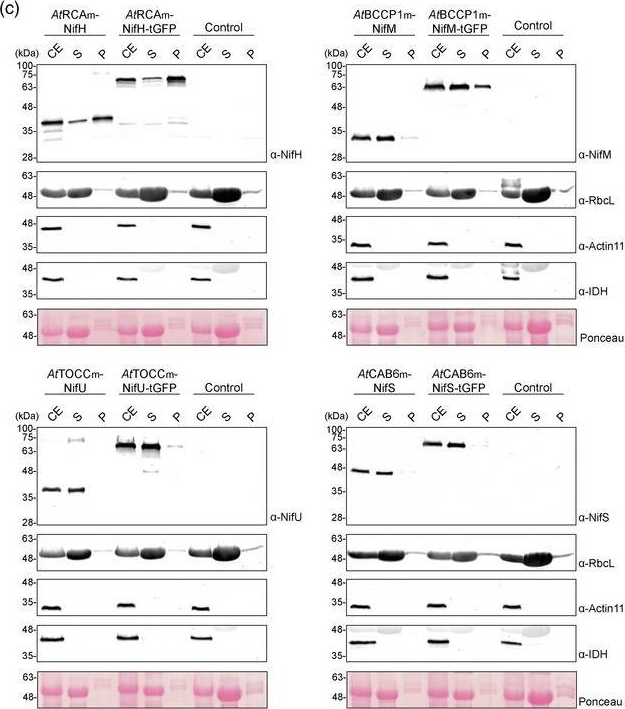
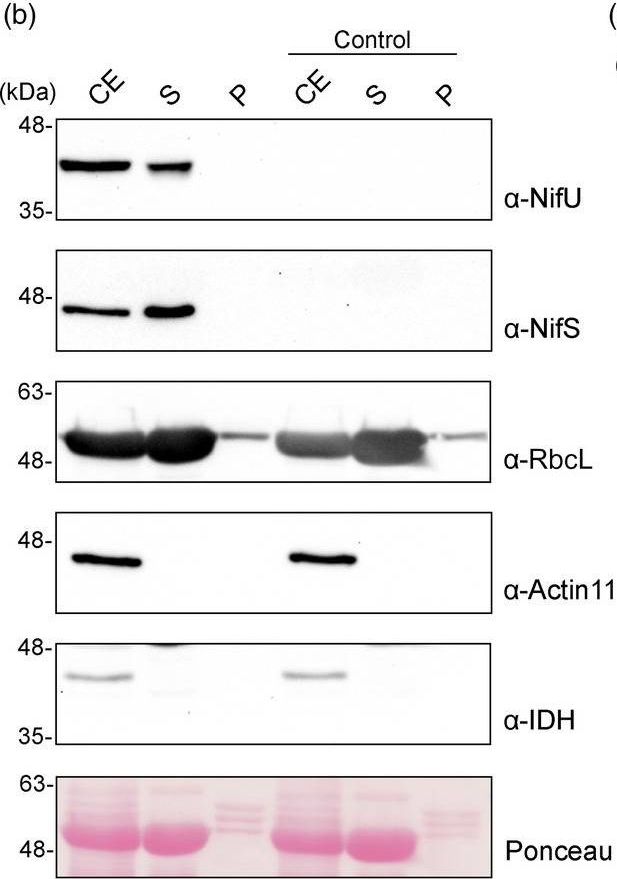
Open PublicationLocalization of stroma?targeted Nif proteins. (a) Schematic representation of the constructs used in N. benthamiana?transient expression assays leading to Nif localization. (b) Confocal microscopy images of Nif proteins fused to tGFP. N.C. corresponds to ‘negative control’, a non?agroinfiltrated leaf. The three columns show the individual signals for tGFP (green, on the left) and chlorophyll autofluorescence (red, in the centre). On the right, overlap of both signals (merge). (c) Chloroplast purification assays. Total proteins extract prior to chloroplast purification (CE). Intact chloroplasts were isolated and then broken to separate the soluble (S) and membrane?associated (P) fractions. Control refers to N. benthamiana plants agroinfiltrated with agrobacterium transformed with an empty vector. Proteins were resolved by SDS?PAGE and detected by Western blot. Expected sizes are Twin?Strep NifH: 35.3 kDa, NifM: 32.3 kDa, NifU: 34.4 kDa, NifS: 44.3 kDa (27 kDa more in the case of tGFP fusions), RbcL: 52.7 kDa, Actin?11: 41.6 kDa and IDH: 39 kDa. Note that the IDH blot was re?probed after striping anti?RuBisco and some signal remains (upper band).
Reactant: Nicotiana tabacum (Common tobacco)
Application: Western Blotting
Pudmed ID: 31985876
Journal: Plant Biotechnol J
Figure Number: 5C
Published Date: 2020-09-01
First Author: Eseverri, Á., López-Torrejón, G., et al.
Impact Factor: 8.66
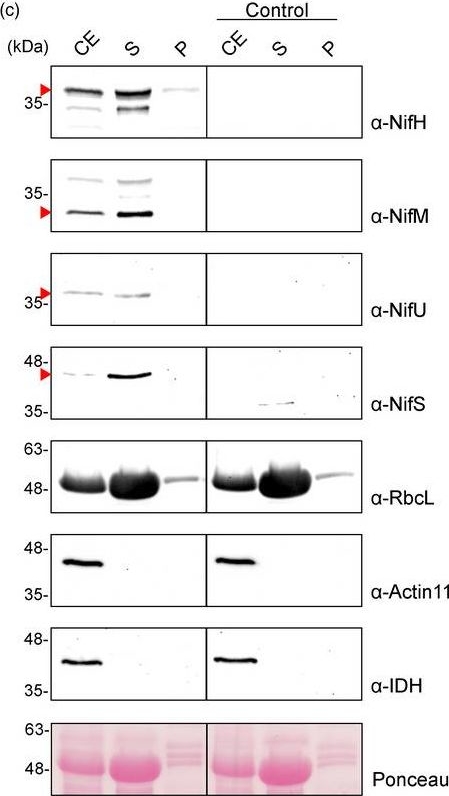
Open PublicationPurification of NifH from chloroplasts of N. benthamiana co?expressing nifH, nifM, nifU and nifS. (a) Schematic representation of the multigenic construct used in N. benthamiana?transient expression assays leading to NifH purification. (b) Western blot analysis of the agroinfiltrated tissue using antibodies targeting Strep?tagged NifH, NifM, NifU and NifS as indicated. Expected sizes are Twin?Strep NifH: 35.3 kDa, NifM: 32.3 kDa, NifU: 34.4 kDa and NifS: 44.3 kDa. (c) Chloroplast purification assays. Total proteins extract prior to chloroplast purification (CE). Intact chloroplasts were isolated and then broken to separate the soluble (S) and membrane?associated (P) fractions. Control refers to N. benthamiana plants agroinfiltrated with agrobacterium transformed with an empty vector. Proteins were resolved by SDS?PAGE and detected by Western blot. Expected sizes are RbcL: 52.7 kDa; Actin?11: 41.6 kDa and IDH: 39 kDa. The triangles in red mark the mature form of the protein. (d) Coomassie staining illustrating the Strep?Tactin purification procedure of A. vinelandii NifH expressed in leaves of N. benthamiana. CFE, cell?free extract (soluble fraction following centrifugation and filtering of disrupted leaf tissue); FT, Strep?Tactin flow?through fraction; W, wash fraction; E, biotin?eluted fraction; Ec, concentrated eluate. (e–f) Western blot analysis of the Strep?Tactin purification procedure. Antibodies targeting Strep?tag (e) or NifH (f) were used for visualization of the purified NifH protein.
Reactant: Nicotiana tabacum (Common tobacco)
Application: Western Blotting
Pudmed ID: 31985876
Journal: Plant Biotechnol J
Figure Number: 6B
Published Date: 2020-09-01
First Author: Eseverri, Á., López-Torrejón, G., et al.
Impact Factor: 8.66
Open PublicationExpression, purification and activity assays of NifU from N. benthamiana chloroplasts. (a) Schematic representation of the construct used in N. benthamiana?transient expression assays leading to NifU purification. pe35s: 35S mosaic cauliflower virus?enhanced promoter; SSU: CTP from the small subunit of RuBisCo; Tnos: nopaline synthase Agrobacterium terminator; p35s: 35S mosaic cauliflower virus promoter. (b) Purification of chloroplasts from NifU and NifS expressing cells. Total proteins extract prior to chloroplast purification (CE). Intact chloroplasts were isolated and then broken to separate the soluble (S) and membrane?associated (P) fractions. Control refers to N. benthamiana plants agroinfiltrated with agrobacterium transformed with an empty vector. Proteins were resolved by SDS?PAGE and detected by Western blot. Expected sizes are Twin?Strep NifU: 36.5 kDa, NifS: 43.7 kDa, RbcL: 52.7 kDa; Actin?11: 41.6 kDa and IDH: 39 kDa. Note that the IDH blot was re?probed after striping anti?RuBisco and some signal remains (upper band). (c) Coomassie staining illustrating the Strep?Tactin purification procedure of A. vinelandii NifU expressed in leaves of N. benthamiana. CFE, cell?free extract (soluble fraction following centrifugation and filtering of disrupted leaf tissue); FT, Strep?Tactin flow?through fraction; W, wash fraction; E, biotin?eluted fraction. (d–f) Western blot analysis of the Strep?Tactin purification procedure. Antibodies targeting Strep?tag (d) or NifU (e) were used for visualization of the purified NifU protein. Antibodies targeting NifS (f) were used to visualize NifS protein co?purifying with NifU. (g) A. vinelandii apo?NifH was activated either by in vitro [4Fe–4S] cluster reconstitution (Rc Apo?NifH) or by incubation with cluster?loaded NifU purified from E. coli (EcNifU) or from N. benthamiana leaves (NbNifU) in 1:1 or 1:2 NifH/NifU molar ratio. Identical reaction mixtures without NifU were used as negative control (Rc C?). Error bars represent mean ± SD (n = 2).
- Additional Information
-
Additional information: Peptide used to elicit this antibody is not conserved in NADPH dependent anzymes, partially conserved across eukaryotic Idh subunits, Some conservation across bacterial which contain the NAD-dependent form of Idh (as opposed to the NADP-dependent form) Additional information (application): Cellular [compartment marker] of mitochondrial matrix
- Background
-
Background: Plant NADH dependent isocitrate dehydrogenase enzyme is located in mitochondrial matrix. This enzyme is classified as an oxidoreductase and its function is to catalyze a reaction in the citric acid cycle, specifically the sequential dehydrogenation and decarboxylation of isocitrate to form a-ketoglutarate. It removes hydrogens from its substrate, isocitrate. In addition to this process, it functions as a decarboxylase, removing a CO2 from the six-carbon substrate to form a five-carbon product mentioned above as a-ketoglutarate. There are two forms of this enzyme NADP+ and NAD+ dependent.
- Product Citations
-
Selected references: Boussardon et al. (2025). The atypical proteome of mitochondria from mature pollen grains. Curr Biol . 2025 Jan 21:S0960-9822(24)01705-6. doi: 10.1016/j.cub.2024.12.037.
Hoa et al. (2024). Proteomic analysis on symbiotic differentiation of mitochondria in soybean nodules. Comparative Study Plant Cell Physiol. 2004 Mar;45(3):300-8. doi: 10.1093/pcp/pch035.
Boussardon et al. (2023). Comparing plastid proteomes points towards a higher plastidial redox turnover in vascular tissues than in mesophyll cells. J Exp Bot. 2023 Apr 7:erad133. doi: 10.1093/jxb/erad133.
Li et al. (2022) The CDC48 complex mediates ubiquitin-dependent degradation of intra-chloroplast proteins in plants. Cell Rep. 2022 Apr 12;39(2):110664. doi: 10.1016/j.celrep.2022.110664. PMID: 35417702.
Kolodziejczak et al. (2018). m-AAA Complexes Are Not Crucial for the Survival of Arabidopsis Under Optimal Growth Conditions Despite Their Importance for Mitochondrial Translation. Plant Cell Physiol. 2018 May 1;59(5):1006-1016. doi: 10.1093/pcp/pcy041.
Rurek et al. (2018). Mitochondrial Biogenesis in Diverse Cauliflower Cultivars under Mild and Severe Drought Involves Impaired Coordination of Transcriptomic and Proteomic Response and Regulation of Various Multifunctional Proteins. Preprints 2018, 2018010276 (doi: 10.20944/preprints201801.0276.v1).
Fujii et al. (2016). The Restorer-of-fertility-like 2 pentatricopeptide repeat protein and RNase P are required for the processing of mitochondrial orf291 RNA in Arabidopsis. Plant J. 2016 Jun;86(6):504-13. doi: 10.1111/tpj.13185.
Yin et al. (2016). Comprehensive Mitochondrial Metabolic Shift during the Critical Node of Seed Ageing in Rice. PLoS One. 2016 Apr 28;11(4):e0148013. doi: 10.1371/journal.pone.0148013. eCollection 2016.
Rurek et al. (2015). Biogenesis of mitochondria in cauliflower (Brassica oleracea var. botrytis) curds subjected to temperature stress and recovery involves regulation of the complexome, respiratory chain activity, organellar translation and ultrastructure. Biochim Biophys Acta. 2015 Jan 21. pii: S0005-2728(15)00016-X. doi: 10.1016/j.bbabio.2015.01.005. - Reviews:
-
Peter Ma | 2019-10-31This antibody works very well for our samples. The membrane proteins from the Arabidopsis roots are subjected to 10% SDS-PAGE and then detected with this antibody (1:5000). The results shows that this antibody is quite specific and sensitive.Michal RUREK | 2014-02-1410 µg of mitochondrial proteins from cauliflower (Brassica oleracea var. botrytis) curds (apical layer) was separated on 12 % SDS-PAGE (Laemmli- type) and blotted 1h to Immobilone P (Millipore) using Sedryt apparatus (Kucharczyk). After blocking (5% milk in PBS-T) for 1 h at RT, blots were incubated in the primary antibody at a dilution of 1: 5000 for 1h at room temperature with agitation. The primary antibody, diluted in 2% milk in PBS-T was reused several times. Secondary, anti-rabbit HRP- linked antibodies were bound at 1/10000 dilution. Single band of 45 kDa was detected using standart GE Healthcare ECL reagents.
Accessories
AS07 212 | Clonality: Polyclonal | Host: Rabbit | Reactivity: A. thaliana, A. palmeri, S. oleracea, di and monocots | Cellular [compartment marker] of mitochondrial outer membrane

AS09 602 | Clonality: Polyclonal | Host: Goat | Reactivity: Rabbit IgG (H&L)

AS09 607 | Clonality: Polyclonal Host: Goat Reactivity: Rabbit IgG (H&L)