1
Anti-Lhcb5 | CP26 chlorophyll a/b-binding protein of plant PSII
AS01 009 | Clonality: Polyclonal | Host: Rabbit | Reactivity: Monocots, dicots and gymnosperms including A. thaliana, C.sinensis, H. vulgare, D. capensis, E. crus-galli, O. sativa, P. sativum, S. oleracea, T. aestivum, Triticale, Z. mays
- Product Info
-
Immunogen: BSA-conjugated synthetic peptide derived from Arabidosis thaliana Lhcb5 protein UniProt: Q9XF89, TAIR: AT4G10340 sequence. This sequence is highly conserved in Lhcb5 proteins from monocots, dicots and conifers but only partial conserved in Physcomitrella patens and Chlamydomonas reinhardtii.
Host: Rabbit Clonality: Polyclonal Purity: Serum Format: Lyophilized Quantity: 50 µl Reconstitution: For reconstitution add 50 µl of sterile water Storage: Store lyophilized/reconstituted at -20°C; once reconstituted make aliquots to avoid repeated freeze-thaw cycles. Please remember to spin the tubes briefly prior to opening them to avoid any losses that might occur from material adhering to the cap or sides of the tube. Tested applications: Immunohistochemistry (IHC), Western blot (WB) Recommended dilution: 1 : 1000 (WB) Expected | apparent MW: 30 | 25-26 kDa for Arabidopsis thaliana
- Reactivity
-
Confirmed reactivity: Arabidopsis thaliana, Camelina sinensis, Echinochloa crus-galli, Hordeum vulgare, Drosera capensis, Oryza sativa, Pisum sativum, Spinacia oleracea, Triticum aestivum, triticale, Zea mays Predicted reactivity: Dicots, Gymnosperms
Not reactive in: No confirmed exceptions from predicted reactivity are currently known - Application Examples
-
.jpg)
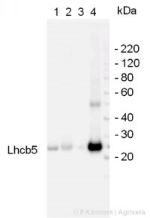
5 µg protein of (1) chloroplasts, (2) thylakoids, and (3) Photosystem I of Arabidopsis thaliana together with (4) BBY (Photosystem II) -particles from Pisum sativum were separated on 4-12% NuPage (Invitrogen) LDS-PAGE and blotted 1h to nitrocellulose. Filters were blocked 1h with 2% low-fat milk powder in TBS-T (0.1% TWEEN 20) and probed with anti-Lhcb5 (AS01 009, 1:1000, 1h) and secondary anti-rabbit (1:20 000, 1 h) antibody (HRP conjugated) in TBS-T containing 2% low fat milk powder. Antibody incubations were followed by washings in TBS-T (15, +5, +5, +5 min). All steps were performed at RT with agitation. Signal was detected with chemiluminescence detection reagent, using a Fuji LAS-3000 CCD (90s, standard sensitivity).
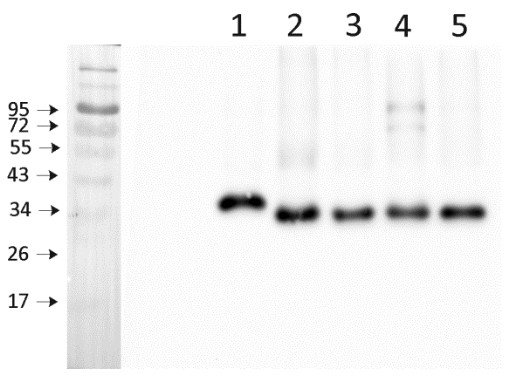
1 µg of chlorophyll from chloroplasts of: Pisum sativum (1), Echinochloa crus-galli, M chloroplasts (2), Echinochloa crus-galli, BS (bundle sheath) chloroplasts (3), Zea mays M chloroplasts (4), Zea mays BS (bundle sheath) chloroplasts (5), extracted with 0.4 M sorbitol, 50 mM Hepes NaOH, pH 7.8, 10 mM NaCl, 5 mM MgCl2 and 2 mM EDTA. Samples were denatured with Laemmli buffer at 75ºC for 5 min and were separated on 12% SDS-PAGE and blotted 30 min to PVDF using wet transfer. Blot was blocked with 5% milk for 2h at room temperature (RT) with agitation. Blot was incubated in the primary antibody at a dilution of 1: 1000 overnight at 4ºC with agitation in 1% milk in TBS-T. The antibody solution was decanted and the blot was washed 4 times for 5 min in TBS-T at RT with agitation. Blot was incubated in secondary antibody (anti-rabbit IgG horse radish peroxidase conjugated, from Agrisera, AS09 602, Lot 2001) diluted to 1:20 000 in 1 % milk in TBS-T for 1h at RT with agitation. The blot was washed 5 times for 5 min in TBS-T and 2 times for 5 min in TBS, and developed for 1 min with 1.25 mM luminol, 0.198 mM coumaric acid and 0.009% H2O2 in 0.1 M Tris- HCl, pH 8.5. Exposure time in ChemiDoc System was 73 seconds.
Courtesy of Dr. Wioleta Wasilewska-Dębowska, Warsaw University, Poland
- Additional Information
-
Additional information: This product can be sold containing ProClin if requested Additional information (application): Protein is processed into mature form (Jansson 1999). - Background
-
Background: Lhcb5 is one of the 3 minor highly conserved chlorophyll a/b-binding proteins associated with Photosystem II in plants and algae. As a part of the inner light-harvesting antenna it has been sugested to regulate (together with Lhcb4 and Lhcb6) the energy flow from the outer LHCII antenna to the PSII reaction center.
- Product Citations
-
Selected references: Wójtowicz et al. (2025). Shrink or expand? Just relax! Bidirectional grana structural dynamics as early light-induced regulator of photosynthesis. New Phytol . 2025 Jun;246(6):2580-2596. doi: 10.1111/nph.70175.
Krupinska et al. (2025). Iron allocation to chloroplast proteins depends on the DNA-binding protein WHIRLY1. Planta. 2025 Jun 17;262(2):32. doi: 10.1007/s00425-025-04736-8.
Ciesielska et al. (2024). S2P2-the chloroplast-located intramembrane protease and its impact on the stoichiometry and functioning of the photosynthetic apparatus of A. thaliana. Front Plant Sci. 2024 Mar 15:15:1372318. doi: 10.3389/fpls.2024.1372318.
Ye et al. (2023). The light-harvesting chlorophyll a/b-binding proteins of photosystem II family members are responsible for temperature sensitivity and leaf color phenotype in albino tea plant. J Adv Res . 2023 Dec 25:S2090-1232(23)00404-6.doi: 10.1016/j.jare.2023.12.017.
Wu et al (2023) Disruption of LEAF LESION MIMIC 4 affects ABA synthesis and ROS accumulation in rice
Ivanov et al. (2022) The decreased PG content of pgp1 inhibits PSI photochemistry and limits reaction center and light-harvesting polypeptide accumulation in response to cold acclimation. Planta 255, 36 (2022). https://doi.org/10.1007/s00425-022-03819-0
Wada et al. (2021) Identification of a Novel Mutation Exacerbated the PSI Photoinhibition in pgr5/pgrl1 Mutants; Caution for Overestimation of the Phenotypes in Arabidopsis pgr5-1 Mutant. Cells. 2021 Oct 26;10(11):2884. doi: 10.3390/cells10112884. PMID: 34831107; PMCID: PMC8616342.
Jeran et al. (2021) The PUB4 E3 Ubiquitin Ligase Is Responsible for the Variegated Phenotype Observed upon Alteration of Chloroplast Protein Homeostasis in Arabidopsis Cotyledons. Genes (Basel). 2021 Sep 6;12(9):1387. doi: 10.3390/genes12091387. PMID: 34573369; PMCID: PMC8464772.
Wojtowicz et al. (2020). Compensation Mechanism of the Photosynthetic Apparatus in Arabidopsis thaliana ch1 Mutants. Int J Mol Sci. 2020 Dec 28;22(1):221. doi: 10.3390/ijms22010221. PMID: 33379339; PMCID: PMC7794896.
Rogowski et al. (2019). Photosynthesis and organization of maize mesophyll and bundle sheath thylakoids of plants grown in various light intensities. Environmental and Experimental Botany Volume 162, June 2019, Pages 72-86.
Krupinska et al. (2019). The nucleoid-associated protein WHIRLY1 is required for the coordinate assembly of plastid and nucleus-encoded proteins during chloroplast development. Planta. 2019 Jan 11. doi: 10.1007/s00425-018-03085-z.
Mao et al. (2018). Comparison on Photosynthesis and Antioxidant Defense Systems in Wheat with Different Ploidy Levels and Octoploid Triticale. Int J Mol Sci. 2018 Oct 2;19(10). pii: E3006. doi: 10.3390/ijms19103006.
Chen et al. (2018). Exogenous melatonin enhances salt stress tolerance in maize seedlings by improving antioxidant and photosynthetic capacity. Physiol Plant. 2018 Apr 6. doi: 10.1111/ppl.12737.
Tyuereva et al. (2017). The absence of chlorophyll b affects lateral mobility of photosynthetic complexes and lipids in grana membranes of Arabidopsis and barley chlorina mutants. Photosynth Res. 2017 Apr 5. doi: 10.1007/s11120-017-0376-9. (Hordeum vulgare, western blot)
Yang-Er Chen et al. (2017). Responses of photosystem II and antioxidative systems to high light and high temperature co-stress in wheat. J. of Exp. Botany, Volume 135, March 2017, Pages 45–55.
Pavlovic et al. (2016). A carnivorous sundew plant prefers protein over chitin as a source of nitrogen from its traps. Plant Physiol Biochem. 2016 Mar 5;104:11-16. doi: 10.1016/j.plaphy.2016.03.008
Sun et al. (2014). Direct energy transfer from the major antenna to the photosystem II core complexes in the absence of minor antennae in liposomes. Biochim Biophys Acta. 2014 Nov 22. pii: S0005-2728(14)00650-1. doi: 10.1016/j.bbabio.2014.11.005
Ido et al. (2014). Cross-Linking Evidence for Multiple Interactions of the PsbP and PsbQ Proteins in a Higher Plant Photosystem II Supercomplex. J Biol Chem. 2014 Jul 18;289(29):20150-7. doi: 0.1074/jbc.M114.574822. Epub 2014 Jun 9.
Zhou et al. (2013) Mutation of the Light-Induced Yellow Leaf 1 Gene, Which Encodes a Geranylgeranyl Reductase, Affects Chlorophyll Biosynthesis and Light Sensitivity in Rice. PLoS ONE 8(9): e75299. doi:10.1371/journal.pone.0075299. - Protocols
-
Agrisera Western Blot protocol and video tutorials
Protocols to work with plant and algal protein extracts
Oxygenic photosynthesis poster by prof. Govindjee and Dr. Shevela
Z-scheme of photosynthetic electron transport by prof. Govindjee and Dr. Björn and Dr. Shevela - Reviews:
-
Shizue Matsubara | 2012-04-04This antibody gave a clear single band with Arabidopsis leaf protein extracts.Maksymilian Zienkiewicz | 2012-01-16This antibody worked efficiently both in A. thaliana and Zea mays plants in suggested working dilutions. There was visible one specific band.
Related products

AS09 602 | Clonality: Polyclonal | Host: Goat | Reactivity: Rabbit IgG (H&L)

AS09 607 | Clonality: Polyclonal Host: Goat Reactivity: Rabbit IgG (H&L)