1
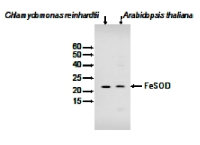
Anti-FeSOD | Fe-dependent superoxide dismutase (chloroplastic)
AS06 125 | Clonality: Polyclonal | Host: Rabbit | Reactivity: A. maritima, A. thaliana, B. juncea, Ch. reinhardtii, D. bardawil, D.salina, M. sativa, Morus spp.,O. sativa, Salicornia sp., S. tuberosum, winter triticale, S. oleracea, Z. mays
- Product Info
-
Immunogen: Overexpressed Chlamydomonas reinhardtii thioredoxine fusion protein A8IGH1, FeSOD excised from a gel piece
Host: Rabbit Clonality: Polyclonal Purity: Serum Format: Lyophilized Quantity: 50 µl Reconstitution: For reconstitution add 50 µl of sterile water Storage: Store lyophilized/reconstituted at -20°C; once reconstituted make aliquots to avoid repeated freeze-thaw cycles. Please remember to spin the tubes briefly prior to opening them to avoid any losses that might occur from material adhering to the cap or sides of the tube. Tested applications: Western blot (WB) Recommended dilution: 1 : 1500-1 : 5000 (WB) Expected | apparent MW: 25 | 22 kDa
- Reactivity
-
Confirmed reactivity: Armeria maritima, Arabidopsis thaliana, Brassica juncea, Chlamydomonas reinhardtii, Dunaliella bardawil, Dunaliella salina, Fremyella diplosiphon, Hordeum vulgare, Medicago sativa, Morus spp., Oryza sativa, Salicornia sp., Solanum tuberosum, winter triticale, Zea mays Predicted reactivity: Algae, Dunaliella salina, Glycine max, Helianthus annuus, Marchantia polymorpha, Nannochloropsis gaditana, Solanum lycopersicum, Physcomitrium patens, Pinus pinaster, Populus balsamifera, Vitis vinifera, Volvox carteri
Species of your interest not listed? Contact usNot reactive in: Spinacia oleracea - Application Examples
-
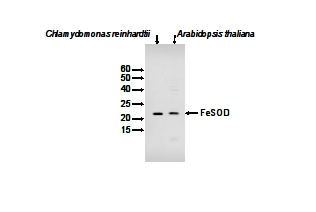
5 µg of stromal protein from (1) Chlamydomonas reinhardtii (left), (2) Arabidopsis thaliana were separated on SDS-PAGE. Primary antibodies have been used in 1: 3000.
- Additional Information
-
Additional information (application): The antibody will detect FeSOD enzyme only in plants grown on low Cu (0.1 μM).Reference: Salah et al (2005) Two P-type ATPases are required for copper delivery in Arabidopsis thaliana chloroplasts. Plant Cell, 17, 1233-1251
Out of three FeSOD isoforms, FeSOD2 and FeSOD3 are not expressed in the roots. In roots of Arabidopsis thaliana, FeSOD1 is detected Takáč et al. (2018)
This product can be sold containing ProClin if requested - Background
-
Background: Antioxidant system works as a defense against oxidative stress. SOD (superoxide dismutase) catalyzes the dismutation of superoxide into oxygen and H,O,. SODs are classified, according to their metal cofactor, as FeSOD, MnSOD, or Cu / ZnSOD. Chloroplasts generally contain Cu/ZnSOD and, in a number of plant species, FeSOD
- Product Citations
-
Selected references: Xing et al. (2026). Transplanting light-dependent reactions for mammalian eye photosynthesis. Cell . 2026 May 15:S0092-8674(26)00469-1. doi: 10.1016/j.cell.2026.04.034.
Holzner et al. (2026). The chloroplast ionome shines light on the dynamics of organellar iron homeostasis. Plant Cell. 2026 Jan 29:koag017. doi: 10.1093/plcell/koag017.
Krupinska et al. (2025). Iron allocation to chloroplast proteins depends on the DNA-binding protein WHIRLY1. Planta. 2025 Jun 17;262(2):32. doi: 10.1007/s00425-025-04736-8.
Grinberg et al (2023) Peptide self-assembly as a strategy for facile immobilization of redox enzymes on carbon electrode.
Fortunato et al. (2022) GUN1 involvement in the redox changes occurring during biogenic retrograde signaling. Plant Science. Volume 320, July 2022, 111265, https://doi.org/10.1016/j.plantsci.2022.111267
Burlacot et al. (2022) Alternative photosynthesis pathways drive the algal CO2-concentrating mechanism. Nature 605, 366–371 (2022). https://doi.org/10.1038/s41586-022-04662-9
Konkolewska et al. (2020). Combined use of companion planting and PGPR for the assisted phytoextraction of trace metals (Zn, Pb, Cd).
Jokel et al. (2020). Elimination of the flavodiiron electron sink facilitates long-term H2 photoproduction in green algae. Biotechnol Biofuels. 2019 Dec 5;12:280. doi: 10.1186/s13068-019-1618-1.
Shull et al. (2019). Anatase TiO2 nanoparticles induce autophagy and chloroplast degradation in thale cress (Arabidopsis thaliana). Environ Sci Technol. 2019 Jul 29. doi: 10.1021/acs.est.9b01648.
Mermod et al. (2019). SQUAMOSA promoter-binding protein-like 7 mediates copper deficiency response in the presence of high nitrogen in Arabidopsis thaliana. Plant Cell Rep. 2019 May 15. doi: 10.1007/s00299-019-02422-0.
Chen et al. (2018). The molecular chaperon AKR2A increases the mulberry chilling-tolerant capacity by maintaining SOD activity and unsaturated fatty acids composition. Sci Rep. 2018 Aug 14;8(1):12120. doi: 10.1038/s41598-018-30379-9.
Bastow et al. (2018). Vacuolar Iron Stores Gated by NRAMP3 and NRAMP4 Are the Primary Source of Iron in Germinating Seeds. Plant Physiol. 2018 Jul;177(3):1267-1276. doi: 10.1104/pp.18.00478.
Hura et al. (2018). Rieske iron-sulfur protein of cytochrome-b6f is involved in plant recovery after drought stress. Rieske iron-sulfur protein of cytochrome-b6f is involved in plant recovery after drought stress.
Balazova et al. (2018). Zinc oxide nanoparticles phytotoxicity on halophyte from genus Salicornia. Plant Physiol Biochem. 2018 Sep;130:30-42. doi: 10.1016/j.plaphy.2018.06.013.
Jokel et al. (2018). Hunting the main player enabling Chlamydomonas reinhardtii growth under fluctuating light. Plant J. 2018 Mar 25. doi: 10.1111/tpj.13897.
Volgusheva et al. (2017). Comparative analyses of H2 photoproduction in magnesium- and sulfur-starved Chlamydomonas reinhardtii cultures. Physiol Plant. 2017 Apr 7. doi: 10.1111/ppl.12576.
Momcilovic et al. (2014). Improved procedure for detection of superoxide dismutase isoforms in potato, Solanum tuberosum L. Acta Physiologiae Plantarum, August 2014, Volume 36, Issue 8, pp 2059-2066.
Dang et al. (2014). Combined Increases in Mitochondrial Cooperation and Oxygen Photoreduction Compensate for Deficiency in Cyclic Electron Flow in Chlamydomonas reinhardtii. Plant Cell. 2014 Jul 2. pii: tpc.114.126375. - Protocols
-
Agrisera Western Blot protocol and video tutorials
Protocols to work with plant and algal protein extracts
Oxygenic photosynthesis poster by prof. Govindjee and Dr. Shevela
Z-scheme of photosynthetic electron transport by prof. Govindjee and Dr. Björn and Dr. Shevela - Reviews:
-
Auroy Pascaline | 2021-04-08I used this Antibody on Chlamydomonas (10µg of protein loading) with dilution 1/1000.Elzbieta Romanowska | 2011-06-30Antibody has been used on Arabidopsis and Armeria leaf extracts with a load per well from 5 to 15 ug and at a dilution of 1: 2000 with alkaline phosphatase conjugated secondary antibody.
Related products
AS09 524 | Clonality: Polyclonal | Host: Rabbit | Reactivity: A. thaliana, A. maritima, A. palmeri, B.napus, B. oleracea, Fragaria x ananassa, N. cataria, N. rtanjensis, O. sativa, P. sativum, R.sativus, Salicornia sp., S. oleracea, S. tuberosum

AS09 602 | Clonality: Polyclonal | Host: Goat | Reactivity: Rabbit IgG (H&L)
AS23 4983 | Clonality: Polyclonal | Host: Rabbit | Reactivity: Arabidopsis thaliana

AS16 3941 | Clonality: Polyclonal | Host: Rabbit | Reactivity: Arabidopsis thaliana

AS06 170 | Clonality: Polyclonal | Host: Rabbit | Reactivity: A.thaliana, A.maritima, B.juncea, D.sanguinalis, E.crus-galli, E.tef, I.pumila, H.vulgare, I. walleriana, L.formosana, N.cataria, M. sativa, N.rtanjensis, O.sativa, P.Sogo Yukidian cultivar V3, P.vulgaris, P.sativum, Populus sp., S.superba, S.lycopersicum, S.tuberosum, Z.mays