1
Anti-PsbE | Alfa subunit of Cytochrome b559 of PSII
AS06 112 | Clonality: Polyclonal | Host: Rabbit | Reactivity: A.thaliana, H. vulgare, N.tabacum, O. sativa, S. oleracea | cellular [compartment marker] of thylakoid membrane
- Product Info
-
Immunogen: KLH-conjugated synthetic peptide chosen from PsbE protein of Arabidopsis thaliana P56779, AtCg00580
Host: Rabbit Clonality: Polyclonal Purity: Serum Format: Lyophilized Quantity: 50 µl Reconstitution: For reconstitution add 50 µl of sterile water Storage: Store lyophilized/reconstituted at -20°C; once reconstituted make aliquots to avoid repeated freeze-thaw cycles. Please remember to spin the tubes briefly prior to opening them to avoid any losses that might occur from material adhering to the cap or sides of the tube. Tested applications: Western blot (WB) Recommended dilution: 1 : 5000 (WB) Expected | apparent MW: 9.25 kDa - Reactivity
-
Confirmed reactivity: Arabidopsis thaliana, Hordeum vulgare, Nicotiana tabacum, Oryza sativa, Spinacia oleracea, Triticum aestivum
Predicted reactivity: Cannabis sativa, Glycne max, Populus alba, Salvia miltiorrhiza, Solanum tuberosum
Species of your interest not listed? Contact usNot reactive in: Chlamydomonas reinhardtii, diatoms, Synechococcus sp. PCC 7942
- Application Examples
-
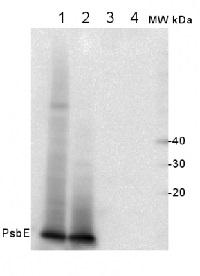
2 µg of total protein from (1) Arabidopsis thaliana leaf, (2) Horderum vulgare leaf ), (3) Chlamydomonas reinhardtii total cell , (4) Synechococcus sp. 7942 total cell were all extracted with Agrisera Protein Extraction Buffer (AS08 300) and separated on 4-12% NuPage (Invitrogen) LDS-PAGE and blotted 1h to PVDF. Blots were blocked immediately following transfer in 2% blocking reagent in 20 mM Tris, 137 mM sodium chloride pH 7.6 with 0.1% (v/v) Tween-20 (TBS-T) for 1h at room temperature with agitation. Blots were incubated with the primary antibody at a dilution of 1: 10 000 for 1h at room temperature with agitation. The antibody solution was decanted and the blot was rinsed briefly twice, then washed once for 15 min and 3 times for 5 min in TBS-T at RT with agitation. Blots were incubated in secondary antibody (anti-rabbit IgG horse radish peroxidase conjugated) diluted to 1:50 000 in 2% blocking solution for 1h/RT with agitation. The blots were washed as above and developed for 5 min with chemiluminescent detection reagent of extreme femtogram range, according to the manufacturers instructions. Images of the blots were obtained using a CCD imager (FluorSMax, Bio-Rad) and Quantity One software (Bio-Rad). Exposure time was 1 second.
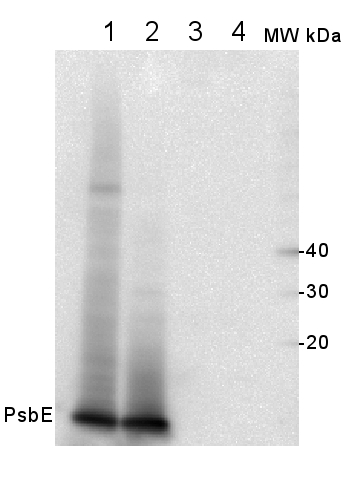
The detection of PsbE proteins was performed using isolated thylakoids from Arabidopsis thaliana and Oryza sativa. Fresh leaf samples (≈ 10 g), 30 min-dark acclimated, were washed and ground in a blender with ice-cold grinding buffer (20 mM Tricine (pH 8.4), containing 0.4 M sorbitol, 5 mM EDTA, 10 mM NaHCO3 and 0.5% BSA). The homogenate was double filtered with two layers of musseline and centrifugated at 4,000 x g for 10 min (4°C). The supernatant was discarded and the pellet was resuspended in an ice-cold resuspending buffer (20 mM Hepes, pH 7.6, 0.3 M sorbitol, 2.5 mM EDTA, 10 mM NaHCO3 and 0.5% BSA). The solution was centrifugated at 4,000 x g for 10 min (4°C), the supernatant was discarded and the pellet was resuspended in a hypotonic buffer (20 mM Hepes, pH 7.6, 2.5 mM EDTA, 10 mM NaHCO3 and 0.5% BSA). The solution was centrifugated (4,000 x g for 10 min, 4°C), the supernatant was discarded and the pellet was resuspended in 1 mL of resuspending buffer. Subsequently, total chlorophyll concentration was determined according to Porra et al. (1989). For the western blot, thylakoid protein extracts were first separated by SDS-PAGE (Laemmli, 1970). Equal volumes (10 mL) of sample containing 4, 2 and 1 µg Chl were electrophoretically transferred to a nitrocellulose membrane (Towbin et al., 1979). Polypeptide detection was performed using specific polyclonal antibodies, anti-PsbE (AS06 112 - AgriserA). Membranes were blocked for 3 hours with 5% non-fat milk in saline Tris-HCl buffer (100 mM Tris-HCl, pH 7.6, 150 mM NaCl), incubated with PsbE antibody overnight and after with HRP-conjugated secondary antibody (AS09 602) by 3 hours. The protein detection was developed using 100 Mm Tris-HCl buffer (pH 7.2) containing 100 mg/mL DAB, 400 mg/mL NiCl2 and 0.04% H2O2, until bands were visualized.
Courtesy of Dr. Ana Karla Moreira Lobo, Laboratory of Plant Metabolism, Federal University of Ceara - Brazil - Additional Information
-
Additional information: Cellular [compartment marker] of thylakoid membrane
This product can be sold containing ProClin if requested.
- Background
-
Background: Cytochrome b559 (Cyt b559) is encoded by the chloroplast genes psbE and psbF and is comprised of two low molecular mass polypeptides, a and h subunits, with molecular masses of 9 and 4 kDa, respectively. The Cyt b559 is closely associated with PSII in all oxygenic photosynthetic organisms. The a and h subunits of the Cyt b559 are components of the minimal PSII reaction center complex that is still capable of primary charge separation In summary, both PsbE and PsbF are essential components for PSII assembly, and they are probably involved in electron transport mechanisms that help to protect PSII from photodamage.
Alternative protein name: PSII reaction center subunit V - Product Citations
-
Selected references: Penzler et al. (2024). A pgr5 suppressor screen uncovers two distinct suppression mechanisms and links cytochrome b6f complex stability to PGR5. Plant Cell. 2024 Mar 27:koae098. doi: 10.1093/plcell/koae098.
Zhao et al. (2024). Psb28 protein is indispensable for stable accumulation of PSII core complexes in Arabidopsis.Plant J. 2024 May 26. doi: 10.1111/tpj.16844.
Hackett et al. (2017). An Organelle RNA Recognition Motif Protein Is Required for Photosystem II Subunit psbF Transcript Editing. Plant Physiol. 2017 Apr;173(4):2278-2293. doi: 10.1104/pp.16.01623.
Yang-Er Chen et al. (2017). Responses of photosystem II and antioxidative systems to high light and high temperature co-stress in wheat. J. of Exp. Botany, Volume 135, March 2017, Pages 45–55.
Nishimura et al. (2016). The N-terminal sequence of the extrinsic PsbP protein modulates the redox potential of Cyt b559 in photosystem II. Sci Rep. 2016 Feb 18;6:21490. doi: 10.1038/srep21490.
Grieco et al. (2015). Light-harvesting II antenna trimers connect energetically the entire photosynthetic machinery - including both photosystems II and I. Biochim Biophys Acta. 2015 Jun-Jul;1847(6-7):607-19. doi: 10.1016/j.bbabio.2015.03.004. Epub 2015 Apr 3.
Hojka et al. (2014). Inducible repression of nuclear-encoded subunits of the cytochrome b6f complex in tobacco reveals an extraordinarily long lifetime of the complex. Plant Physiol. 2014 Jun 24. pii: pp.114.243741. - Protocols
-
Agrisera Western Blot protocol and video tutorials
Protocols to work with plant and algal protein extracts - Reviews:
-
This product doesn't have any reviews.
Related products

AS09 602 | Clonality: Polyclonal | Host: Goat | Reactivity: Rabbit IgG (H&L)
AS04 038 | Clonality: Polyclonal | Host: Rabbit | Reactivity: [global antibody] for higher plants, Physcomitrella patens, algae, cyanobacteria, diatoms
Benefits of using this antibody
AS09 522 | Clonality: Polyclonal | Host: Rabbit | Reactivity: Arabidopsis thaliana, Pinus strobus L.
AS05 084 | Clonality: Polyclonal | Host: Rabbit | Reactivity: [global antibody] for higher plants, algae, liverwort, cyanobacteria, diatoms | cellular [compartment marker] of thylakoid membrane
Benefits of using this antibody
AS06 146 | Clonality: Polyclonal | Host: Rabbit | Reactivity: [global antibody] for A. thaliana, Anabaena 7120, D. brightwellii, H. vulgare, C.reinhardtii, C. zofingiensis, L. corniculatus, N. tabacum, O. sativa, P. sativum, P. vulgaris, P. tricornutum, T. pratense, S. alba, Synechococcus sp. PCC 7942, Synechocystis sp. PCC6803, T. guillardii, T. pseudonana, Triticale, U. prolifera, Z. mays
Benefits of using this antibody